Introduction to EEG
EEG Signal Processing
인간의 뇌의 신경 활동은 태아 발육의 17주 ~ 23주 사이에 시작된다. 이러한 초기 단계 혹은 평생을 통해, 뇌에 의해 생성된 전기 신호는 뇌기능뿐만 아니라 전신의 상태(Whole body)도 나타낼 수 있다고 가정해보자. 이러한 가정은 고급 디지털 신호 처리 방법을 인간 피험자의 뇌에서 측정된 뇌파 (EEG) 신호에 적용해볼 수 있는데, 이를 통해 논문의 다음 장에서 논해보기로 한다. 이 논문은 뇌 활동의 생리적 측면에 대해 언급할 생각은 없지만, 원래 Source의 본질, 실제 패턴 및 반드시 해결되어야 할 매체의 특성과 관련된 몇 가지 문제가 있다. 매체는 소위 신호원 (signal source)이라고 불리는 뉴런으로부터의 경로를 정의하며, 센서는 Source의 혼합물 형태가 측정되는 전극에 연결된다. 그러나 뇌의 신경 기능 및 신경생리학적 성질과 신호 생성 및 그 기록의 기본 메커니즘은 뇌 질환 및 관련 질병의 탐지, 진단 및 치료를 위해 이러한 신호를 다루는 사람들에게 중요하다. 뇌파 측정(EEG)에 대한 간단한 역사기록을 살펴보자.
1.1 History
Carlo Matteucci (1811-1868)와 Emil Du Bois-Reymond (1818-1896)는 검류계(galvanometer)를 사용하여 근육 신경에서 방출된 전기 신호를 등록한 최초의 사람들이었으며 신경생리학(neurophysiology) 개념을 수립했다.
그러나 Hermann Von Helmholtz [3]에 의해 도입된 활동 전류(action current)의 개념은 근육 수축 중에 생기는 음의 변동을 분명히 확인했다.
영국 리버풀 출신의 과학자 리차드 카튼 (Richard Caton, 1842-1926)은 검류계를 사용하여 피검자의 두피(scalp) 위에 두 개의 전극을 놓았으며, 그로인해 1875 년에 전기 신호의 형태로 뇌 활동을 기록했다.
그 이후로 electro-(뇌의 전기적 활동 등록), encephalo-(머리에서 신호를 방출하는 것을 말함), 그리기 또는 쓰기를 의미하는 gram(또는 graphy)이 결합되어 이후,
EEG라는 용어는 뇌의 전기적 신경 활동을 나타내기 위해 사용되었다.
Fritsch (1838-1927)와 Hitzig (1838-1907)는 인간 대뇌가 전기적으로 자극 될 수 있음을 발견했다.
Vasili Yakovlevich Danilevsky (1852-1939)는 Caton의 연구에 이어 1877 년에 뇌의생리학 연구에서 박사 학위 논문을 마쳤다.
이 연구에서, 그는 뇌의 전기 자극뿐만 아니라 동물 두뇌에서의 자발적인 전기 활동을 조사했다.
다른 동물 종의 시각 피질에서 관찰 된 (대)뇌 전기 활동은 Ernst Fleischl von Marxow (1845-1891)에 의해 보고되었다.
나폴레옹 사이 불 스키 (Napoleon Cybulski, 1854-1919)는 전기 자극으로 인한 개에 간질 발작의 뇌파 증거를 제공했다.
간질 발작(epileptic seizure)과 비정상적인 전기적 방전(Discharge, 흐름)의 연관성(association)에 대한 개념은 Kaufman [5]에 의해 표현되었다.
러시아의 생리학자인 Pravidch-Neminsky (1879-1952)는 뇌에서 뇌파(EEG)를 기록했으며, 1912 년에 경질(dura)이라 불리는 뇌파와 손상되지 않은 개의 두개골(skull)을 기록했다.
개는 정상적인 상태에서 12-14cycle/s의 리듬을 보았는데, 이는 질식 하에서 느려지고 나중에 "electrocerebrogram" 이라고 불렀다.
인간의 뇌파 신호의 존재를 발견 한 사람은 한스 버거 (Hans Berger, 1873-1941)였다. 그는 1920 년에 인간 뇌파(EEG) 연구를 시작 했다[6]. 버거는 거의 모든 뇌파전위기록자로 잘 알려져 있으며 1910 년에 현 검류계 작업을 시작하여 작은 에델 맨 모델로 전환하고 1924 년 이후 큰 에델 맨 모델로 전환했다. 1926 년 Berger는 더 강력한 Siemens 더블 코일 검류계 (130μV / cm의 감도를 달성)을 사용하기 시작했다 [7]. 1929 년에 사진논문에 1 ~ 3 분간의 인간 뇌파 기록에 관한 그의 첫 번째 보고서가 있었다. 이 기록에서는 전두엽 - 후두부 리드(?)가 있는 1 채널 바이폴라 방식만 사용했다. EEG의 기록은 1924 년에 인기를 얻었다. 1929 년 Berger의 첫 번째 보고서에는 이 장의 뒷부분에서 설명하는 대로 EEG 신호의 주요 구성 요소인 알파 리듬과 알파 차단 응답이 포함되었다.
1930 년대에 수면 스핀들의 첫 번째 뇌파 기록은 버거 (Berger)에 의해 수행되었다. 그 후 그는 저산소증이 인간의 뇌에 미치는 영향, 여러 가지 확산되고 국소화 된 뇌 질환의 특성을 보고하고 간질성 발작을 암시한다고 보고했다. 이 시간 동안 Berlin-Buch에 설립되어 Kornmuller가 이끄는 다른 그룹은 뇌파를 보다 정확하게 기록했다. Berger는 뇌 국소화(대뇌피질에서 담당하는 기능이 특정 부위에 국한되어 존재하는 현상), 특히 뇌종양의 국소화에도 관심이 있었고, 정신 활동과 뇌파 신호의 변화 사이에 약간의 상관관계를 발견했다.
베를린의 그룹 Toennies (1902-1970)은 뇌 전위를 기록하는 최초의 생물학적 증폭기를 제작했다. EEG를 기록하기 위한 차동 증폭기는 1932 년 록펠러 재단에 의해 제조되었다.
멀티 채널 기록의 중요성과 더 넓은 뇌 영역을 커버하기 위해 다수의 전극을 사용하는 것은 Kornmuller [10]에 의해 인식되었다. 간질 발작에 초점을 맞춘 첫 번째 EEG 연구와 첫 번째 데모는 Fischer andL¨owenbach [11-13]에 의해 제시되었다.
영국에서는 W. Gray Walter가 임상 뇌파 검사의 선구자가 되었다. 그는 느린 뇌 활동 (델타 파)의 초점을 발견했는데, 이는 뇌 이상 진단에 엄청난 임상적 관심을 불러 일으켰다.
브뤼셀에서는 프레드릭 브레머 (Fredric Bremer, 1892-1982)가 각성 상태에 대한 구심성 신호의 영향을 발견했다.
EEG 관련 연구 활동은 1934 년경 북미에서 시작되었다.
올해 Hallowell Davis는 훌륭한 알파 리듬을 보여주었다.
이 시기에 워싱턴 주 세인트루이스 대학 (St Louis University)의 그룹이 음전선 오실로스코프를 사용하여 말초 신경 잠재력 연구에 나섰다.
인간 뇌파에 대한 작업은 1930 년대 보스턴의 하버드 (Harvard)와 아이오와 대학 (University of Iowa)에서 시작되었다.
간질 발작 장애의 영역이 가장 큰 효과를 발휘 했으므로 Fredric Gibbs가 개발 한 간질 발작 연구는 이 기간 동안 EEG의 주된 작업이었다.
간질은 역사적으로 뇌파가 출현하기 전/후에 2 가지[15]로 나눌 수 있다.
Gibbs와 Lennox는 picrotoxin에 대한 그의 연구와 인간 간질에 동물에서 대뇌 피질 뇌파에 미치는 영향을 기반으로 피셔의 아이디어를 적용하였다.
Berger [16]은 부전 마비를 가진 환자에서 경련 발작의 추정 및 경련 발작 동안 발작의 EEG 방전의 몇 가지 예를 보여 주었다.
북미에서 뇌파 검사의 다른 훌륭한 개척자인 할로 엘과 폴린 데이비스 (Pauline Davis)는 인간의 수면 중에 뇌파의 본질에 대한 최초의 연구자였다.
A. L. Loomis, E. N. Harvey 및 G. A. Hobart는 인간 수면 뇌파 패턴과 수면 단계를 수학적으로 연구 한 최초의 사람이었다.
맥길 대학 (McGill University)에서 H. Jasper는 기초 및 임상 간질에서 틈새를 발견하기 전에 관련 행동 장애를 연구했다[17].
미국 뇌파 학회는 1947 년에 설립되어 그동안 영국 런던에서 제 1 회 국제 뇌파 회의가 개최되었다. 독일 뇌파 검사는 여전히 베를린에 한정되어 있었지만,
일본은 뇌파 리듬의 연구자 인 모토카와 연구로 주목 받고있다 [18].
그 동안, 신경 생리학은 해부학 적 방법을 통해 시상 피질의 관계를 보여 주었다. 이것은 중추성 간질의 개념의 발달로 이어졌다 [19].
1950 년대 내내 EEG에 대한 연구가 여러 곳에서 진행되었고, 이 기간 동안 간질 병을 제거하기 위한 외과 수술이 인기를 얻었다.
간질과 인간 두뇌의 기능적 해부학(Penfiled and Jasper)이라는 책이 출판되었다.
이 기간 동안 미세 전극이 발명되었다. 그 발명품들은 텅스텐 또는 유리와 같은 금속으로 만들어졌으며, 염화칼륨과 같은 전해질로 채워진 원통모양이었고 직경은 3 μm 미만이었다.
인간의 심도 뇌파 검사는 Mayer와 Hayne (1948)가 이식 한 뇌내 전극을 통해 처음으로 얻어졌다.
세포 내 미세 전극 기술의 발명은 이 방법을 혁명적으로 바꾸었고 1952 년 Brock 등의 척수에서 사용했고 1961 년 Phillips의 피질에서 사용되었다.
뇌파 신호 분석은 뇌파 측정 초기에 시작했다.
Dierg (1932)가 도와주어 Berger는 1950 년대에 급속하게 발전된 EEG 시퀀스에 푸리에 분석을 적용했다.
EEGs를 이용한 수면 장애의 분석은 시카고 대학의 Kleitman의 연구를 통해 1950 년대에 시작되었다.
1960 년대에는 만삭아와 조기 신생아의 뇌파 분석이 개발되기 시작했다 [20].
정신 질환 모니터링에 흔히 사용되는 evoked potentials (EP), 특히 비주얼 EP의 조사가 1970 년대에 진행되었다.
그러나 EEG의 역사는 1300 년대 초부터 시작하여 수많은 신경 학적 및 생리 학적 이상 발견, 인식, 진단, 치료에 대한 임상 실험 및 계산 연구의 일상 발전을 가져온 인간의 중추 신경계 (CNS)의 나머지 부분 사이에 존재한다.
요즘 EEG는 완전히 전산화 된 시스템을 사용하여 침습적이며 비침습적으로 기록된다.
EEG 기계는 많은 신호 처리 도구, 섬세하고 정확한 측정 전극 및 몇 시간의 장기간 기록을 위한 충분한 메모리가 장착되어 있으며
EEG 또는 MEG (magnetoencephalogram) 기계는 기능적 자기 공명 영상 (fMRI)과 같은 다른 신경 영상 시스템과 통합 될 수 있다.
매우 섬세한 바늘 형 전극은 또한 피질 (electrocortiogram)에서 EEG를 기록하기 위해 사용될 수 있으며,
따라서 두개골에 의해 유도되는 감쇠 및 비선형 효과를 피할 수 있다.
인간 두뇌 내에서의 신경 활동의 본질은 다음 장에 규명 될 것이다.
------------------------------------------------------------------------------------------------------------------------------------------------------
논문 1.2 Neural Activities에 들어가기 앞서...
◆Neuron이란 전기 및 화학적인 신호를 서로 주고 받으며 정보를 주고 받는 세포들이다. 하지만 이렇게 유명한 neuron말고도 신경계에는 glia에 속하는 세포들도 존재한다.
◆glia라는 이름은 그리스어로 아교를 뜻하는 glue에서 유래되었는데 '접착제' 처럼 neuron들을 둘러싸는 역할을 수행하는 세포이다.
또한, neuron에서 주변에 있는 neuron으로 신호를 보낼 수 있도록 하는 세포 내 돌기로써 axon을 감싸는 glia의 종류도 존재한다.
◆즉, axon은 축삭돌기로써, 전류와 비슷한 형태로 다른 뉴런으로 신호를 전달하는 기능이 있다.
◆dendron은 그리스어로 나무를 의미하며, neutron에서 나뭇가지 형태를 하고 있는 부분의 명칭이 dendrite인 이유이다. neuron이 자극을 받았을 때 전기적인 신호는 dendrite를 통해 접수되어(input) 핵과 소기관이 들어있는 cell body를 통과하여 axon으로 진행한다.
이 때 cell body에서 axon으로 넘어가는 부분을 axon hillock이라고 부른다.
◆이 때 언급한 cell body, 즉 신경세포체는 여러 neuron으로부터 전달되는 외부자극에 대한 판정, 다른 neuron으로 신호를 전달할지 말지를 결정한다.
◆마지막으로, 그림에 나와있지는 않지만 synapse는 A 뉴런의 축삭돌기 말단과 다음 B 뉴런의 수상돌기의 연결부위를 말하며, 이 시냅스는 얇은 막 형태라 다른 뉴런의 축삭돌기로부터 받는 신호를 어느정도의 세기로 전달할지를 결정한다.
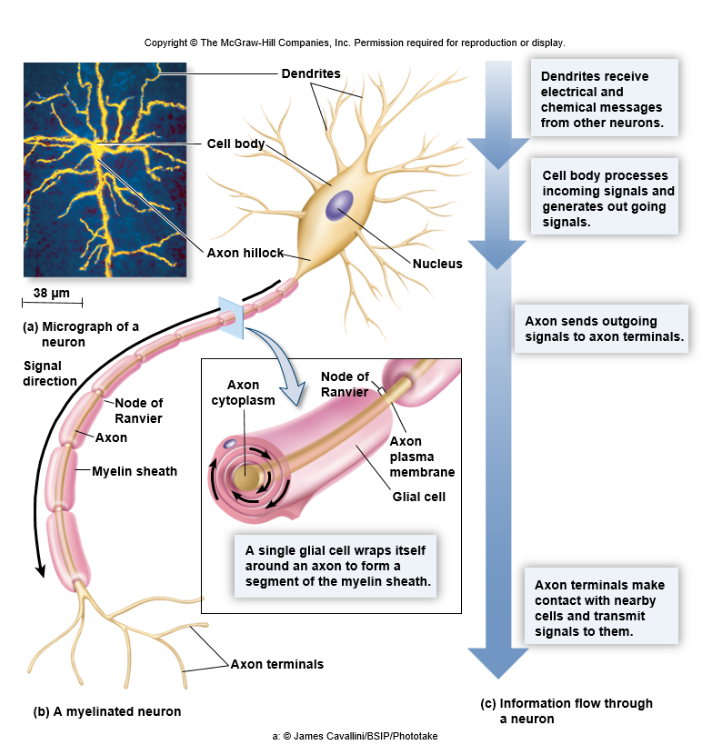 [배경지식] Neuron의 구조
[배경지식] Neuron의 구조◆neuron에서는 세포막을 기준으로 안쪽과 바깥쪽에 있는 전하량의 차이가 존재한다. 이를 membrane potential이라 부르는데,
◆세포가 신호를 전달하고 있지 않는 평상시의 막전위를 휴지전위, resting potential(-70mv)이라 부른다.
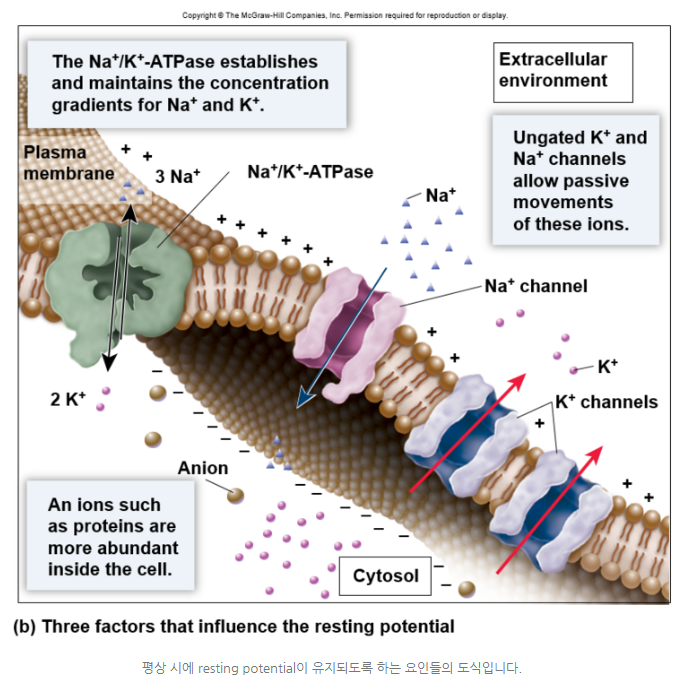 [배경지식]막전위: 휴지전위 (Membrane Potential: Resting Potential)
[배경지식]막전위: 휴지전위 (Membrane Potential: Resting Potential) ◆neuron은 이온의 이동을 통해서 막전위를 변화시키고, 이러한 변화가 neuron에서 다음 neuron으로 이동하면서 신호가 전달된다.
◆막전위가 변화하는 양상에는 탈분극(depolarization)과 과분극(hyperpolarization)이 있는데, 특히 -70mV인 휴지 전위(resting potential)가 -70mV 정도였다.
membrane potential이 음의 방향으로 커질 때(예를 들어 -80mV) hyperpolarization이라고 하고, 이러한 polarization이 약해질 때를 depolarization이라고 한다(-50mV와 같이 0에 더 가까워질 때!).
◆활동전위(action potential)를 논하지 전에 차등전위에 대해서 얘기해보자, 자극이 주어졌을 때, 그 자극의 강도에 따라서 depolarization과 hyperpolarization의 정도가 달라지는 경우를 차등전위(graded potential)이라고 부르는데 이러한 graded potential는 dendrite나 cell body에서 국소적으로 발생하는 현상이다.
화학적 또는 전기적 자극에 의해서 이온채널이 열려서 depolarization이나 hyperpolarization이 일어나는 건 다들 알 것이다.
이 때 이온채널이 열리는 개수에 따라서 이러한 전위 변화의 정도가 달라지는데, 아래 그림에서는 붉은 선이 상대적으로 강한 자극, 검은 선이 상대적으로 약한 자극을 받았을 때의 전위 변화를 보여준다.
그렇게 때문에graded potential 충분히 크면 활동 전위(action potential)로 넘어간다!
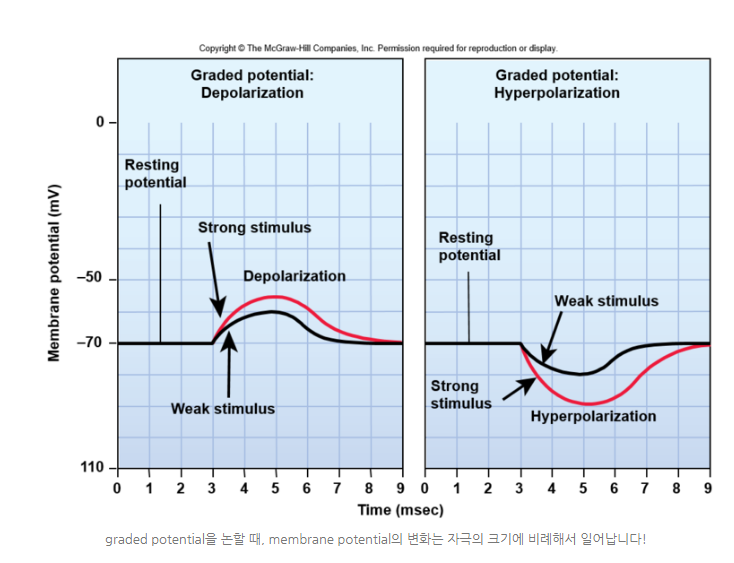 [배경지식]차등 전위 (Graded Potential)
[배경지식]차등 전위 (Graded Potential) ◆graded potential이 threshold potential 값을 넘게 되면 action potential이 된다.
고등학교 때 실무율의 원리라고 들어봤을텐데, 엄청나게 대단한 용어인 것처럼 들리지만 영어로 보면 실망잼
◆all or none law
◆threshold potential을 넘기면 action potential(all)이지만 못 넘기면 그대로 슈르르륵 사라지는 것이다(none).
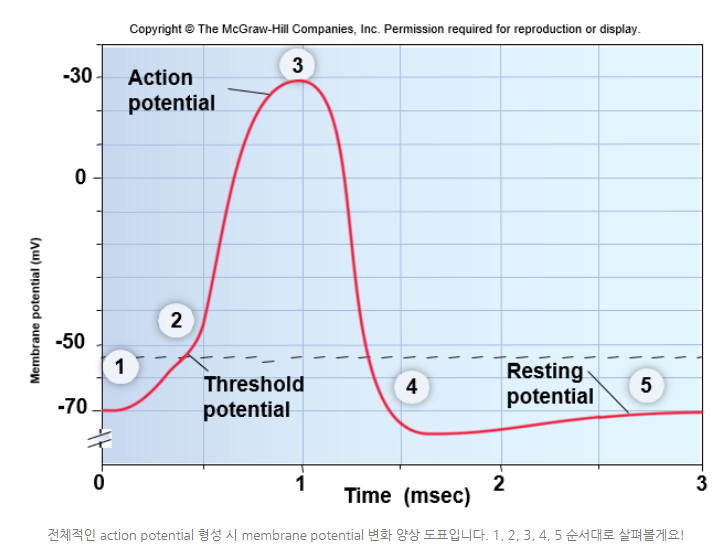 [배경지식]활동 전위 (Action Potential)
[배경지식]활동 전위 (Action Potential) ◆다음으로 EPSP와 IPSP에 관한 내용이다.
간단히 말하자면, 이들은 시냅스들의 신호전달방법을 알리는 것으로써 이들 각각은 기억 방식 중 하나이다.
우선, EPSP[Excitatory Post-Synaptic Potential]는 흥분성 시냅스 후 전위이며, 시냅스 뒷부분의 신경은 탈분극을 일으키는 시냅스로써, 이를 전달하는 물질은 주로 Na+이다.
반면, IPSP[Inhibitory Post Synaptic Potential]는 억제적 시냅스 후 전위이며, 뒷부분의 신경세포에서는 신경이 억제되는 효과를 보이는데 후부의 신경세포는 과분극을 일으키며 K+가 유출되거나 CI-를 유입시키는 방법으로 작용한다.
이쯤 했으면 배경지식은 어느정도 쌓였기 때문에 다음 논문을 보는데 무리없을 것이다.
------------------------------------------------------------------------------------------------------------------------------------------------------
1.2 Neural Activities
중추신경계(CNS)는 일반적으로 신경 사이에 위치하는 신경 세포(nerve cells)와 신경교세포(glia cells)로 구성되며, 각 신경 세포는 축삭, 수상 돌기 및 세포체로 구성된다.
신경 세포는 자극에 반응하고 장거리 사이에서 정보를 전달한다. 신경세포체는 단일 핵을 가지고 있으며 신경 세포 대사의 대부분을 포함한다. 특히 단백질 합성과 관련이 있다. 세포체에서 생성된 단백질은 신경의 다른 부위로 전달된다.
축색 돌기(axon)는 전기적 충격(impulse)을 전달하는 긴 원통기둥 모양이며 척추동물에서 수 미터의 길이가 될 수 있다.(기린의 축색 돌기는 머리에서 척추 끝까지 간다.)
인간의 경우, 길이는 1mm에서 1M 이상이 될 수 있다. 세포의 말단에 단백질을 전달하기 위한 축색 수송 시스템이 존재하며 수송 시스템에는 튜브린 레일로 이동하는 '분자 모터'가 있다.
수상 돌기(dendrite)는 다른 세포의 축색 돌기 또는 수상 돌기에 연결되어 다른 신경에서 충동을 받거나 다른 신경에 신호를 전달한다. 인간의 두뇌에서 각 신경은 주로 돌기 연결을 통해 약 10,000 개의 다른 신경과 연결된다 .
중추 신경계의 활동은 주로 축삭과 수상 돌기의 접합부(=시냅스) 또는 세포의 수상 돌기 (dendrites)와 수상 돌기 (dendrites) 사이에서 전달되는 시냅스 전류와 관련이 있다. 마이너스 극성을 갖는 60-70 mV의 전위가 세포체의 막 아래에 기록 될 수 있다. 이 전위는 시냅스 활동의 변화에 따라 변한다.
활동 전위가 흥분성(excitatory) 시냅스에서 끝나는 섬유를 따라 이동하면 흥분성 시냅스 후 전위Excitatory Post-Synaptic Potential(EPSP)가 다음 뉴런에서 발생한다.(뉴런 간 시냅스에서 기억 모형의 매듭(node)간 연결에서 이전 단위의 흥분이 다음 단위의 흥분을 일으키는 혹은 강화하는 관계를 말함) 두 개의 활동 전위가 짧은 거리에 걸쳐 동일한 섬유를 따라 이동하는 경우 막 전위의 특정 임계값에 도달하는 시냅스 후위 뉴런에 활동 전위를 생성하는 EPSP가 합계에 도달한다.
섬유가 억제(inhibitory) 시냅스에서 끝나면, 과분극이 일어나며, 억제 시냅스 후 전위 Inhibitory Post Synaptic Potential(IPSP)을 나타낸다 [21,22](학습은 기억 내 매듭(node)간 연결강도가 변화하는 것을 말하며 연결강도가 nagative로 변화되고 이를 억제적이라 말함.) IPSP 생성 후, 신경 세포에서 양이온이 넘치거나 신경 세포로 음이온이 유입되는데, 이 흐름은 궁극적으로 신경 세포막을 따라 전위 변화를 일으킨다.
그림 1.1은 위의 활동을 개략적으로 보여준다.
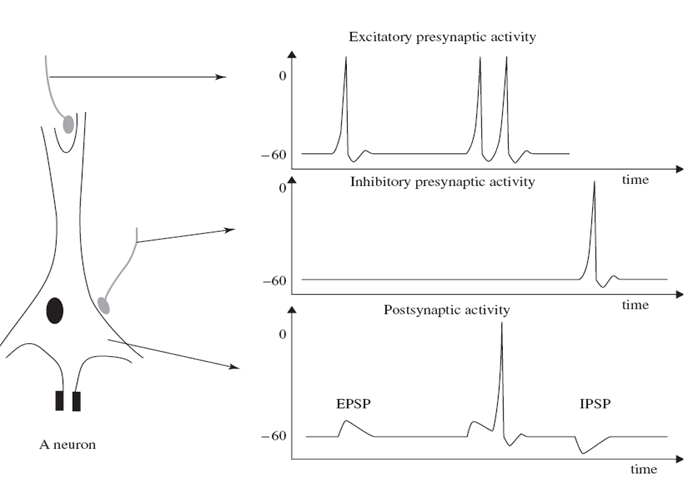 그림 1.1 시냅스 활성화 동안, 신경 세포 전위 변화와 전류 흐름은 세포 내 미세 전극의 수단에 의해 기록한다. 흥분성 및 억제성 시냅스 섬유의 활동 전위는 각각 시냅스 후부에 있는 뉴런에서 EPSP 및 IPSP를 유도한다.
그림 1.1 시냅스 활성화 동안, 신경 세포 전위 변화와 전류 흐름은 세포 내 미세 전극의 수단에 의해 기록한다. 흥분성 및 억제성 시냅스 섬유의 활동 전위는 각각 시냅스 후부에 있는 뉴런에서 EPSP 및 IPSP를 유도한다.1차 막 전류 (transmembranous current)는 세포 내/외 공간에서 세포막을 따라 2차 inonal(?) 전류를 생성한다. 세포 외 공간을 통해 흐르는 이러한 전류의 부분은 field 전위의 발생을 직접적으로 담당하며, 일반적으로 100 Hz 미만의 주파수를 갖는이 필드 전위는 신호 평균에 변화가 없는 경우 EEG라고 하며 평균 신호에 느린 드리프트가 있으면 실제 EEG 신호를 마스킹 할 수 있다. EEG와 DC 전위의 조합은 종종 발작 (pentylenetetrazol에 의해 유도 됨), 고칼슘혈증 및 질식과 같은 뇌의 일부 이상에서 관찰된다 [23].
다음 장은 활동전위의 본질에 초점을 맞춘다.
------------------------------------------------------------------------------------------------------------------------------------------------------
논문 1.3 Action Potential에 들어가기 앞서...
과연 action potential의 정체는 무엇인가?..
지금부터 어떤 과정으로 Action Potential이 형성되는지 살펴보자
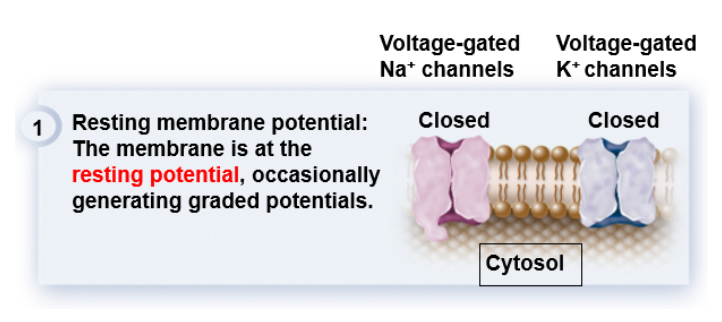 [배경지식] voltage-gated channels이 Na+와 K+ 둘 모두 닫혀있음
[배경지식] voltage-gated channels이 Na+와 K+ 둘 모두 닫혀있음1) 평소에 뉴런의 세포막이 resting potential을 유지하고 있을 때는 voltage-gated channels이 Na+와 K+ 둘 다에 대해서 모두 닫혀있다.
또는, 가끔씩 자극이 오면 graded potential을 형성하기 위해 잠깐 열렸다 닫히기도 한다.
◆여기서 voltage-gated channels란 뭘까???
channel이라고해서 마냥 열려만 있는 것은 아니다. 어떤 특정 조건을 만족해야지 gate가 열리는데, 그 기준에 따라 ligand-gated channel과 voltage-gated channel, 마지막으로 mechanosensitive channel이 있다. 또한 열려있다고 해서 모든 물질이 그 통로를 통과할 수 있는 것은 아니다.
예를 들어 aquaporin이라는 channel은 막을 통해 물의 이동을 가능하게 하지만 물 이외의 것들은 통과 못한다.
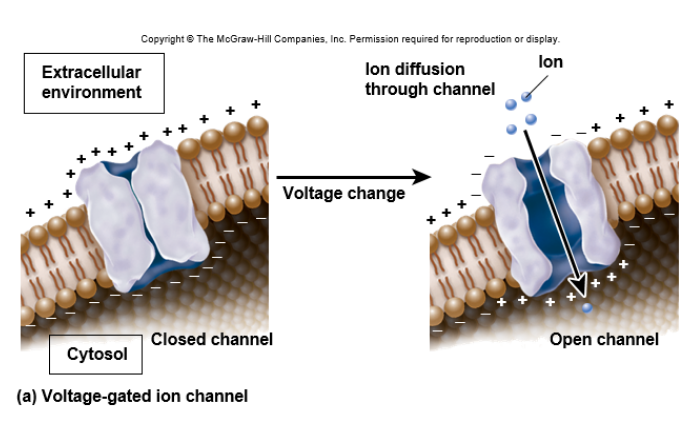 [배경지식] voltage-gated channel
[배경지식] voltage-gated channel◆ligand-gated는 호르몬이나 신경전달물질 같이 어떤 신호를 전달하는 분자(ligand)가 channel에 붙으면 gate가 열리는 방식인 반면,
◆ligand-gated처럼 channel에 직접적으로 분자가 결합하지 않는 경우인, voltage-gated는 이름대로 세포막을 사이로 한 전위의 변화에 따라 개폐가 결정된다. 신경세포의 Na+와 K+ channel이 유명한 예시이다.
◆차등전위graded potential로 인한 depolarization이 -50이나 -55mV 정도에 도달하면 역치를 넘어 action potential이 발생한다. voltage-gated Na+ channel이 열려서 뉴런 안쪽으로 Na+ 이온들이 쏟아져 들어오게 되며 결국 depolarization는 더 심해질 것이다. 그래서 총괄적인 membrane potential 도표에서 3번에 해당하는 상태까지 가게 될것이다.
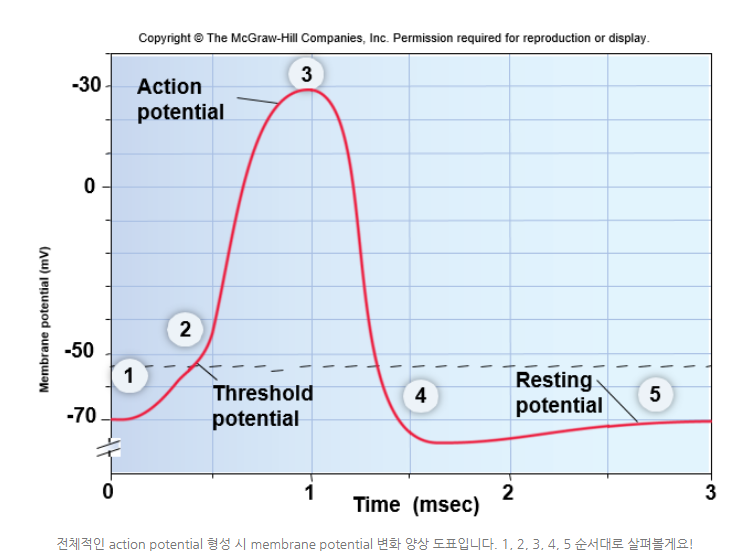 [배경지식] 총괄적인 membrane potential 도표
[배경지식] 총괄적인 membrane potential 도표3) 도표 상 3이 위치한 값은 +50mV 정도다.
여기까지 도달하고 나면 활짝 열렸던 voltage-gated Na+ channel들이 비활성되고, K+ channel들이 열린다!!
전위 값의 변화에 반응하는 속도가 Na+ channel에 비해서 K+ channel이 느리고, 이로 인해 이렇게 어찌보면 복잡한 막전위의 변화 단계들이 만들어진 것이다.
Na+ channel이 비활성화 된다는 건 channel이 닫히지는 않았지만 임시로(?) 닫혔다는 말이다.
Na+ channel이 +50mV나 되는 membrane potential에 노출되면 inactivation gate라는게 돌출되면서 channel의 통로를 막아버리기 때문이다.
그림에서 살펴보면 channel 전체가 완전히 닫히지는 않았지만 좌우지간 이온이 이동할 길이 막혀버린걸 확인할 수 있다.
한편 K+는 평상시에 뉴런 안쪽에서의 농도가 높으므로 channel이 열리면 뉴런 바깥쪽으로 이동할 것이다.
그럼 다시 뉴런의 막전위membrane potential가 음의 값을 갖는 방향으로 이동하게 된다.
inactivation gate가 막은 Na+ channel은 완전히 닫힌건 아니기 때문에 외부 자극이 오든 말든 무반응(불응기)이다.
이렇게 방금 action potential을 일으킨 뉴런의 경우 다시 action potential이 발생할 가능성이 일시적으로 차단되어 있는 상태이기 때문에 자극이 한쪽 방향으로만 전달되는 효과를 가져온다.
난 이미 활동전위action potential 전달을 방금 했으니 아직 활동전위action potential를 못 형성한 dendrite에서 axon terminal쪽으로 다음 뉴런에게 이 자극을 전달해주겠어! 하는 것이다.
 [배경지식] 3
[배경지식] 3K+ 이온들이 와장창 빠져나갔으니 막전위membrane potential가 다시 휴지전위resting potential에 가까워진다. 4단계에 도달하면 아까 비활성화 됐던 Na+ channel들이 이젠 완전히 닫히게 된다.
그 와중에 K+ channel들은 계속 열려있다보니 휴지전위resting potential에 가까워진 정도가 아니라 더 낮아진다.
저번에 네른스트 방정식을 이용해서 각 이온 별로 평형 전위를 구했었죠? K+는 그 값이 -90mV 정도였습니다.
K+가 와장창 이동하니까 K+에 의해서만 전위가 형성되는 듯한 상황이 펼쳐지면서, 그 전위 값이 -90mV에 더 가까워지는 것이고, 이를 hyperpolarization이라고 부르게 된다.
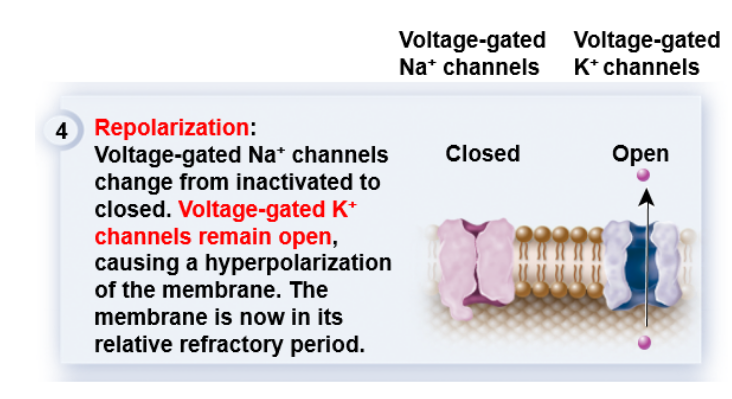 [배경지식] 4
[배경지식] 45) 자~! 이제 마지막 단계이다. resting potential로 되돌아가야 할 때가 왔다.
Na+와 K+에 대한 channel이 둘 다 닫히고, Na+/K+ pump가 다시 열일을 시작하면, 우리와 친숙한 -70mV라는 값의 막전위membrane potential가 등장한다.
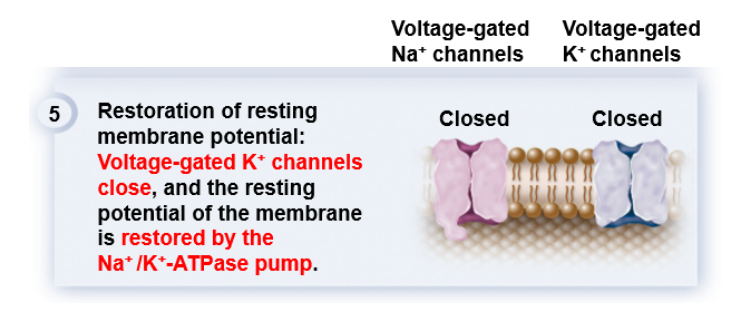 [배경지식] 5
[배경지식] 5지금까지, 주구장창 등장한 주인공들은 Na+와 K+ channel들이며, 이 두 존재가 체내의 모든 신호전달의 주역이라는 사실은 반대로 이 둘에게 무슨 문제가 생기면 생명체에게 치명적이라고 해석할 수도 있다.
따라서 많은 신경독들이 Na+와 K+ channel의 활동을 방해하는 방식을 택한 이유가 되기도 한다. 복어독인 테트로도톡신(tetrodotoxin)이나 보톡스의 재료인 보툴리닌(botulinin)이 다 신경독이다.
이쯤 했으면 배경지식은 어느정도 쌓였기 때문에 다음 논문을 보는데 무리없을 것이다.
------------------------------------------------------------------------------------------------------------------------------------------------------
1.3 Action Potentials
신경에 의해 전달되는 정보를 활동 전위Action Potential(AP)이라고 한다. AP는 뉴런 막을 가로 질러 이온이 교환 됨으로써 발생하며, AP는 축삭을 따라 전달되는 막전위의 일시적인 변화이다. 그것은 일반적으로 세포체에서 시작되어 한 방향으로 이동한다. 막 전위는 탈분극(더 긍정적이 됨)하여 스파이크를 일으킨다. 스파이크의 피크 후에 막이 재분극한다. (보다 음적으로). 전위는 휴지 전위보다 음수가되고 정상으로 돌아 간다. 대부분의 신경의 활동 전위는 5 ~ 10 m/s초 동안 지속된다.
그림 1.2는 예제 AP를 보여준다.
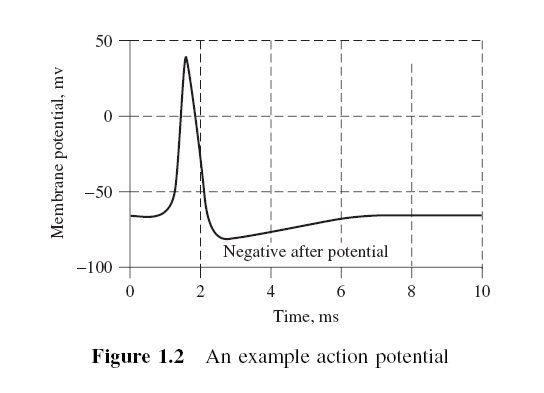 그림 1.2 Action Potential 예제
그림 1.2 Action Potential 예제활동 전위의 전도 속도는 1 ~ 100 m / s입니다. AP는 다양한 유형의 자극에 의해 시작됩니다. 감각 신경은 화학, 빛, 전기, 압력, 촉각, 스트레칭과 같은 다양한 유형의 자극에 반응합니다. 반면에, CNS 내의 신경 (뇌 및 척수)은 대부분 시냅스에서의 화학적 활성에 의해 자극을 받는다. AP를 시작하려면 자극이 임계 값 이상이어야 합니다. 매우 약한 자극은 작은 국소 전기 교란을 일으키지 만 전송 된 AP는 생성하지 않습니다. 자극 강도가 임계 값을 초과하면 행동 잠재력이 나타나 신경을 따라 이동한다.
AP의 스파이크는 주로 Na (나트륨) 채널의 열림에 기인합니다. Na 펌프는 Na와 K (칼륨) 이온의 그라데이션을 생성합니다. 둘 다 활동 잠재력을 생산하는 데 사용됩니다. Na는 셀 바깥 쪽이 높고 내부가 낮습니다. 흥분성 세포는 멤브레인 전압 (전압 게이팅 채널)에 반응하여 게이트가 열리고 닫히는 특수 Na 및 K 채널을 가지고 있습니다. Na 채널의 게이트를 열면 Na가 셀로 들어가서 양의 전하를 띠게됩니다. 이것은 멤브레인 전위를 양극으로 만들고 (탈극) 스파이크를 만듭니다. 그림 1.3은 거대한 오징어에 대한 활동 전위가 진화하는 동안의 과정의 단계를 보여준다. 인간의 경우 AP의 진폭은 약 -60 mV와 10 mV 사이다. 이 과정에서 [24] :
I. 신경 세포의 수상 돌기가 자극을 받으면 Na + 채널이 열립니다. 개구부가 내부 전위를 -70mV에서 -55mV까지 끌어 올리는데 충분하다면 프로세스는 계속됩니다.
II. 동작 임계 값에 도달하면 추가 Na + 채널 (때로는 전압 제어 채널이라고 함)이 열립니다. Na + 유입은 세포막 내부를 약 + 30 mV까지 증가시킵니다. 이 과정을 탈분극이라고합니다.
III. 그런 다음 Na + 채널이 닫히고 K + 채널이 열립니다. K + 채널이 열리는 속도가 훨씬 느리기 때문에, 탈분극을 완료 할 시간이 있습니다. Na +와 K + 채널을 동시에 열어 놓으면 시스템이 중립성을 갖게되고 활동 전위가 생성되는 것을 막을 수 있습니다.
IV. K + 채널을 열면 멤브레인은 휴식 잠재력으로 다시 재분극하기 시작합니다.
V. 재분극은 일반적으로 잔류 전위를 약 -90 mV 수준까지 오버 슈트 (overshoot)한다. 이것은 과분극 (hyperpolarization)이라고 불리며 비생산적인 것처럼 보이지만 사실 정보의 전송에서는 중요합니다. 과분극은이 시간 동안 뉴런이 다른 자극을받지 못하게하거나 새로운 자극에 대한 임계 값을 적어도 올립니다. 과분극의 중요성의 일부는 이미 축삭을 보내어 자극이 반대 방향으로 다른 활동 전위를 유발하는 것을 방지하는 데 있습니다. 즉, 과분극은 신호가 한 방향으로 진행되는 것을 보증합니다.
VI. 과분극 후, Na + / K + 펌프는 막을 -70 mV의 안정 상태로 되돌려 놓습니다.
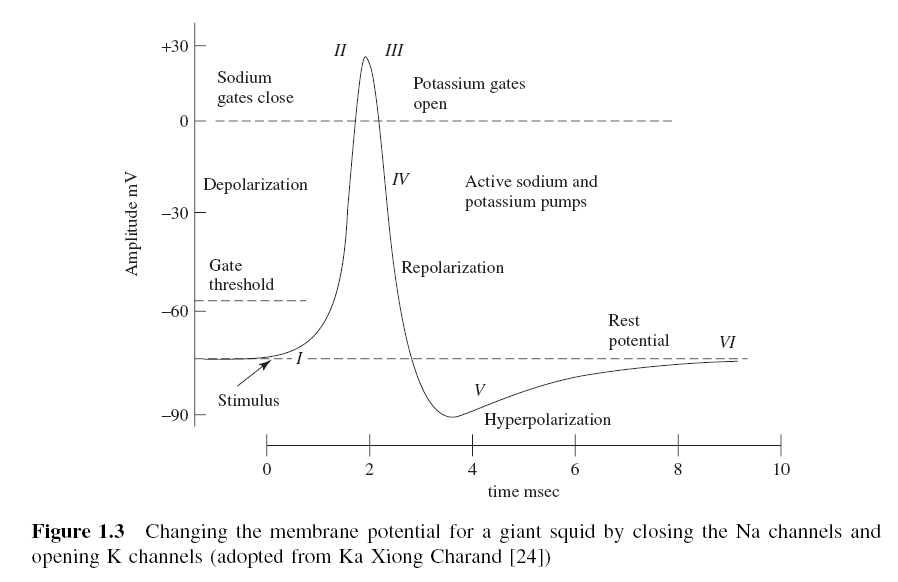 그림 1.3 Action Potential 과정
그림 1.3 Action Potential 과정신경은 다른 자극이 나타 내기 전에 약 2m/s가 필요하다. 이 시간 동안 AP를 생성 할 수 없기 때문에 이것을 불응 기간이라고 한다.
다음에 뇌파 신호의 생성에 대해 설명할 것이다.
References
[1] Caton, R., ‘The electric currents of the brain’, Br. Med. J., 2, 1875, 278.
[2] Walter, W. G., ‘Slow potential waves in the human brain associated with expectancy, attention and decision’, Arch. Psychiat. Nervenkr., 206, 1964, 309–322.
[3] Cobb, M., ‘Exorcizing the animal spirits: Jan Swammerdam on nerve function’, Neuroscience, 3, 2002, 395–400.
[4] Danilevsky, V. Y., ‘Investigation into the physiology of the brain’ [in Russian], Doctoral Thesis, University of Kharkov, 1877, Zit. Nach: Brazier MAB; A history of Neurophysiology in the 19th Century, New York: Raven; 1988, 208.
[5] Brazier, M. A. B., A History of the Electrical Activity of the Brain; The First Half-Century, Macmillan, New York, 1961.
[6] Massimo, A., ‘In Memoriam Pierre Gloor (1923–2003): an appreciation’, Epilepsia, 45(7), July 2004, 882.
[7] Grass, A. M., and Gibbs, F. A., ‘A Fourier transform of the electroencephalogram’, J. Neurophysiol., 1, 1938, 521–526.
[8] Haas, L. F., ‘Hans Berger (1873–1941), Richard Caton (1842–1926), and electroencephalography’, J. Neurol. Neurosurg. Psychiat., 74, 2003, 9.
[9] Spear, J. H., ‘Cumulative change in scientific production: research technologies and the structuring of new knowledge’, Perspectives on Sci., 12(1), 2004, 55–85.
[10] Shipton, H. W., ‘EEG analysis: a history and prospectus’, Annual Rev., Univ. of Iowa, USA, 1975, 1–15.
[11] Fischer, M. H., ‘Elektrobiologische Auswirkungen von Krampfgiften am Zentralnervensystem’, Med. Klin., 29, 1933, 15–19.
[12] Fischer, M. H., and Lowenbach, H., ‘Aktionsstrome des Zentralnervensystems unter der Einwirkung von Krampfgiften, 1. Mitteilung Strychnin und Pikrotoxin’, Arch. F. Exp. Pathol. und Pharmakol., 174, 1934, 357–382.
[13] Kornmuller, A. E., ‘Der Mechanismus des Epileptischen Anfalles auf Grund Bioelektrischer Untersuchungen am Zentralnervensystem’, Fortschr. Neurol. Psychiatry, 7, 1935, 391–400; 414–432.
[14] Bremer, F., ‘Cerveau isole’ et physiologie du sommeil’, C.R. Soc. Biol. (Paris), 118, 1935, 1235–1241.
[15] Niedermeyer, E., ‘Historical aspects’, Chapter 1, Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn., Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 1–14.
[16] Berger, H., ‘Uber das Elektrenkephalogramm des Menschen’, Arch. Psychiatr. Nervenkr., 87, 1929, 527–580.
[17] Jasper, H., ‘Report of committee on methods of clinical exam in EEG’, Electroencephalogr. Clin. Neurophysiol., 10, 1958, 370–375.
[18] Motokawa, K., ‘Electroencephalogram of man in the generalization and differentiation of condition reflexes’, Tohoku J. Expl. Medicine, 50, 1949, 225.
[19] Niedermeyer, E., ‘Common generalized epilepsy. The so-called idiopathic or centrencephalic epilepsy’, Eur. Neurol., 9(3), 1973, 133–156.
[20] Aserinsky, E., and Kleitman, N., ‘Regularly occurring periods of eye motility, and concomitant phenomena, during sleep’, Science, 118, 1953, 273–274.
[21] Speckmann, E.-J., and Elger, C. E., ‘Introduction to the neurophysiological basis of the EEG and DC potentials’, in Electroencephalography Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999.
[22] Shepherd, G. M., The Synaptic Organization of the Brain, Oxford University Press, London, 1974.
[23] Caspers, H., Speckmann E.-J., and Lehmenk¨uhler, A., ‘DC potentials of the cerebral cortex, seizure activity and changes in gas pressures’, Rev. Physiol., Biochem. Pharmacol., 106, 1986, 127–176.
[24] Ka Xiong Charand, http://hyperphysics.phy-astr.gsu.edu/hbase/biology/actpot.html.
[25] Attwood, H. L., and MacKay, W. A., Essentials of Neurophysiology, B. C. Decker, Hamilton, Canada, 1989.
[26] Nunez, P. L., Neocortical Dynamics and Human EEG Rhythms, Oxford University Press, New York, 1995.
[27] Teplan, M., ‘Fundamentals of EEG measurements’, Measmt Sci. Rev., 2(2), 2002.
[28] Bickford, R. D., ‘Electroencephalography’, in Encyclopedia of Neuroscience, Ed. G. Adelman,Birkhauser, Cambridge (USA), 1987, 371–373.
[29] Sterman, M. B., MacDonald, L. R., and Stone, R. K., ‘Biofeedback training of sensorimotor EEG in man and its effect on epilepsy’, Epilepsia, 15, 1974, 395–416.
[30] Ashwal, S., and Rust, R., ‘Child neurology in the 20th century’, Pedia. Res., 53, 2003, 345–361.
[31] Niedermeyer, E., ‘The normal EEG of the waking adult’, Chapter 10, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 174–188.
[32] Pfurtscheller, G., Flotzinger, D., and Neuper, C., ‘Differentiation between finger, toe and tongue movement in man based on 40 Hz EEG’, Electroencephalogr. Clin. Neurophysiol., 90, 1994, 456–460.
[33] Adrian, E. D., and Mattews, B. H. C., ‘The Berger rhythm, potential changes from the occipital lob in man’, Brain, 57, 1934, 345–359.
[34] Trabka, J., ‘High frequency components in brain waves’, Electroencephalogr. Clin. Neurophysiol., 14, 1963, 453–464.
[35] Cobb, W. A., Guiloff, R. J., and Cast, J., ‘Breach rhythm: the EEG related to skull defects’, Electroencephalogr. Clin. Neurophysiol., 47, 1979, 251–271.
[36] Silbert, P. L., Radhakrishnan, K., Johnson, J., and Class, D. W., ‘The significance of the phi rhythm’, Electroencephalogr. Clin. Neurophysiol., 95, 1995, 71–76.
[37] Roldan, E., Lepicovska, V., Dostalek, C., and Hrudova, L., ‘Mu-like EEG rhythm generation in the course of Hatha-yogi exercises’, Electroencephalogr. Clin. Neurophysiol., 52, 1981, 13.
[38] IFSECN, ‘A glossary of terms commonly used by clinical electroencephalographers’, Electroencephalogr. Clin. Neurophysiol., 37, 1974, 538–548.
[39] O’Leary, J. L., and Goldring, S., Science and Epilepsy, Raven Press, New York, 1976, pp. 19–152.
[40] Gotman, J., Ives, J. R., and Gloor, R., ‘Automatic recognition of interictal epileptic activity in prolonged EEG recordings’, Electroencephalogr. Clin. Neurophysiol., 46, 1979, 510–520.
[41] ‘Effects of electrode placement’, http://www.focused-technology.com/electrod.htm, California.
[42] Collura, T., A Guide to Electrode Selection, Location, and Application for EEG Biofeedback, Ohio, Brain- Master Technologies, Inc. 1998.
[43] Nayak, D., Valentin, A., Alarcon, G., Seoane, J. J. G., Brunnhuber, F., Juler, J., Polkey, C. E., and Binnie, C. D., ‘Characteristics of scalp electrical fields associated with deep medial temporal epileptiform discharges’, Clin. Neurophysiol., 115, 2004, 1423–1435.
[44] Barrett, G., Blumhardt, L., Halliday, L., Halliday, A. M., and Kriss, A., ‘A paradox in the lateralization of the visual evoked responses’, Nature, 261, 1976, 253–255.
[45] Halliday, A. M., Evoked potentials in neurological disorders’, in Event-Related Brain Potentials in Man, Eds E. Calloway, P. Tueting, and S. H. Coslow, Academic Press, New York, 1978, 197–210.
[46] Sharbrough, F. W., ‘Nonspecific abnormal EEG patterns’, Chapter. 12, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes Da Silva, 4th edn., Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999.
[47] Bancaud, J., Hecaen, H., and Lairy, G. C., ‘Modification de la reactivite EEG, troubles des functions symboliques et troubles con fusionels dans les lesions hemispherigues localisees’, Electroencephalogr. Clin. Neurophysiol., 7, 1955, 179.
[48] Westmoreland, B., and Klass, D., ‘Asymmetrical attention of alpha activity with arithmetical attention’, Electroencephalogr. Clin. Neurophysiol., 31, 1971, 634–635.
[49] Cobb, W., ‘EEG interpretation in clinical medicine’, Part B, in Handbook of Electroencephalography and Clinical Neurophysiology, Ed. A. Remond, Amsterdam, Vol. 11, Elsevier, 1976.
[50] Hess, R., ‘Brain tumors and other space occupying processing’, Part C, in Handbook of Electroencephalography and Clinical Neurophysiology, Ed. A. Remond, Amsterdam, Vol. 14, Elsevier, 1975.
[51] Klass, D., and Daly, D. (Eds), Current Practice of Clinical Electroencephalography, 1st edn. Raven Press, 1979.
[52] Van Sweden, B., Wauquier, A., and Niedermeyer, E., ‘Normal aging and transient cognitive disorders in the elderly’, Chapter 18, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 340–348.
[53] America Psychiatric Association, Committee on Nomenclature and Statistics, Diagnostic and Statistical Manual of Mental Disorder: DSM-IV , 4th edn., American Psychiatric Association, Washington, DC, 1994.
[54] Brenner, R. P., ‘EEG and dementia’, Chapter 19, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn., Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 349–359.
[55] Neufeld, M. Y., Bluman, S., Aitkin, I., Parmet, Y., and Korczyn, A. D., ‘EEG frequency analysis in demented and nondemented Parkinsonian patients’, Dementia, 5, 1994, 23–28.
[56] Niedermeyer, E., ‘Abnormal EEG patterns: epileptic and paroxysmal’, Chapter 13, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 235–260.
[57] Hughes, J. R., and Gruener, G. T., ‘Small sharp spikes revisited: further data on this controversial pattern’, Electroencephalogr. Clin. Neurophysiol., 15, 1984, 208–213.
[58] Hecker, A., Kocher, R., Ladewig, D., and Scollo-Lavizzari, G., ‘Das Minature-Spike-Wave’, Das EEG Labor, 1, 1999 51–56.
[59] Geiger, L. R., and Harner, R. N., ‘EEG patterns at the time of focal seizure onset’, Arch. Neurol., 35, 1978, 276–286.
[60] Gastaut, H., and Broughton, R., Epileptic Seizure, Charles C. Thomas, Springfield, Illinois, 1972.
[61] Oller-Daurella, L., and Oller-Ferrer-Vidal, L., Atlas de Crisis Epilepticas, Geigy Division Farmaceut, Spain, Barcelona 1977.
[62] Niedermeyer, E., ‘Nonepileptic Attacks’, Chapter 28, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 586–594.
[63] Creutzfeldt, H. G., ‘Uber eine Eigenartige Herdformige Erkrankung des Zentralnervensystems’, Z. Ges. Neurol. Psychiatr., 57, 1968, 1, Quoted after W. R. Kirschbaum, 1920.
[64] Jakob, A., ‘Uber Eigenartige Erkrankung des Zentralnervensystems mit Bemerkenswerten Anatomischen Befunden (Spastistische Pseudosklerose, Encephalomyelopathie mit Disseminerten Degenerationsbeschwerden)’, Deutsch. Z. Nervenheilk , 70, 1968, 132, Quoted after W. R. Kirschbaum, 1921.
[65] Niedermeyer, E., ‘Epileptic seizure disorders’, Chapter 27, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 476–585.
[66] Small, J. G., ‘Psychiatric disorders and EEG’, Chapter 30, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th ed., Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 235–260.
[67] Marosi, E., Harmony, T., Sanchez, L., Becker, J., Bernal, J., Reyes, A., Diaz de Leon, A. E., Rodriguez, M., and Fernandez, T., ‘Maturation of the coherence of EEG activity in normal and learning disabled children’, Electroencephalogr. Clin. Neurophysiol. 83, 1992, 350–357.
[68] Linden, M., Habib, T., and Radojevic, V., ‘A controlled study of the effects of EEG biofeedback on cognition and behavior of children with attention deficit disorder and learning disabilities’, Biofeedback Self Regul., 21(1), 1996, pp. 35–49.
[69] Hermens, D. F., Soei, E. X., Clarke, S. D., Kohn, M. R., Gordon, E., and Williams, L. M., ‘Resting EEG theta activity predicts cognitive performance in attention-deficit hyperactivity disorder’, Pediatr. Neurol., 32(4), 2005, 248–256.
[70] Swartwood, J. N., Swartwood, M. O., Lubar, J. F., and Timmermann, D. L., ‘EEG differences in ADHDcombined type during baseline and cognitive tasks’, Pediatr. Neurol., 28(3), 2003, 199–204.
[71] Clarke, A. R., Barry, R. J., McCarthy, R., and Selikowitz, M., ‘EEG analysis of children with attentiondeficit/ hyperactivity disorder and comorbid reading disabilities’, J. Learn. Disabil., 35(3), 2002, 276–285.
[72] Yordanova, J., Heinrich, H., Kolev, V., and Rothenberger, A., ‘Increased event-related theta activity as a psychophysiological marker of comorbidity in children with tics and attention-deficit/hyperactivity disorders’, Neuroimage, 32(2), 2006, 940–955.
[73] Jacobson, S., and Jerrier, H., ‘EEG in delirium’, Semin. Clin. Neuropsychiat., 5(2), 2000, 86–92.
[74] Onoe, S., and Nishigaki, T., ‘EEG spectral analysis in children with febrile delirium’, Brain Devel., 26(8), 2004, 513–518.
[75] Brunovsky, M., Matousek, M., Edman, A., Cervena, K., and Krajca, V., ‘Objective assessment of the degree of dementia by means of EEG’, Neuropsychobiology, 48(1), 2003, 19–26.
[76] Koenig, T., Prichep, L., Dierks, T., Hubl, D., Wahlund, L. O., John, E. R., and Jelic, V., ‘Decreased EEG synchronization in Alzheimer’s disease and mild cognitive impairment’, Neurobiol. Aging, 26(2), 2005, 165–171.
[77] Babiloni, C., Binetti, G., Cassetta, E., Dal Forno, G., Del Percio, C., Ferreri, F., Ferri, R., Frisoni, G., Hirata, K., Lanuzza, B., Miniussi, C., Moretti, D. V., Nobili, F., Rodriguez, G., Romani, G. L., Salinari, S., and Rossini, P. M., ‘Sources of cortical rhythms change as a function of cognitive impairment in pathological aging: a multicenter study’, Clin. Neurophysiol., 117(2), 2006, 252–268.
[78] Bauer, G., and Bauer, R., ‘EEG, drug effects, and central nervous system poisoning’, Chapter 35, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn, Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 671–691.
[79] Naidu, S. and Niedermeyer, E., ‘Degenerative disorders of the central nervous system’, Chapter 20, in Electroencephalography, Basic Principles, Clinical Applications, and Related Fields, Eds E. Niedermeyer and F. Lopes da Silva, 4th edn., Lippincott, Williams and Wilkins, Philadelphia, Pennsylvania, 1999, 360–382.
Paper List
Today's Word
이 논문을 이해하기 위해서 중요한 단어들 수록
- Neurophysiology 신경생리학
- galvanometer 검류계
- cerebral (대)뇌의
- Physiology 생리학
- cortex 피질
- epileptic 간질의
- discharges 방전(흐름)
- skull 두개골
- electroencephalographer 뇌파전위기록자
- fronto-occipital 전두엽과 후두엽
- hypoxia 저산소증
- tumours 종양
- foci 초점
- afferent 구심성
- vigilance 각성
- cathode ray 음극선
- electroencephalography 뇌파검사
- electrodes 전극
- peripheral nerve potential 말초신경전위
- picrotoxin 피크로톡신물에 약간 녹는 백색결정으로, 쓴맛나는 유독물질
- tetrodotoxin 북어독
- petit mal 소간질1초~수초 간 지속되는 의식상실 및 근육경련의 발작
- thalamocortical 사상피질
- microelectrodes 미소전극
- electrolytes 전해질
- potassium chloride 염화칼륨
- spinal 척수
- dura 개의 뇌 막,(개의 뇌 외부에는 경질(dura-meter), 지주막, 피아 라는 3개의 막으로 덮여있다.)
- Seizure/spikes/manifestation 발작
- paroxysmal 발작성의
- motor seizure 중첩성 간질
- Asphysiz 질식
- Abnormality 이상, 장애
- Central Nervous System(CNS) 중추신경계
- Potential 전위(전기적 위치 에너지로써 어떤 양전하를 기준점부터 어느 점까지 이동시키기 위한 일의 양이 바로 이 위치에너지)
- cell body/soma 세포체
- Dendrite 수상돌기, 가지돌기
- Axon 축삭(돌기), 축색(돌기)
- Glia cells 신경교세포
- membrane potential 막전위뉴런에서는 세포막 기준으로 왼쪽과 바깥쪽에 있는 전하량의 차이가 존재하는데 이를 막전위라 부름
- resting potential 휴지전위막전위에서 세포를 전달하지 않는 평상시엔 휴지상태임
- Hyperpolarization 과분극막전위가 음의방향으로 커질 때, 과분극이라 함
- Threshold potential 역치전위
- Action potential 활동전위
- Hypercapnia 고칼슘혈증
- EPSP 흥분성 시냅스 후 전위
- IPSP 억제적 시냅스 후 전위
- Cation 양이온 Anions 음이온